



200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过





(点击图片查看学习列表)
哌拉西林他唑巴坦与依替米星联合治疗CAP,为何仍未好转?
案例
老年男性,慢性病程,急性起病。
诊断
细菌性肺炎、高血压、冠心病。
诊疗经过
患者两周前出现发热,体温最高39 ℃,伴畏寒、寒战,伴全身肌肉关节酸痛,伴鼻塞、流涕、打喷嚏,伴咳嗽、咳痰,无喘息、呼吸困难、胸闷、咳血、呕吐等不适,遂就诊入院。入院后血常规化验结果:白细胞:11.45*109/L,中性粒细胞百分比:80 %,快速C反应蛋白:123 mg/L。肝肾功能正常。考虑细菌性肺炎,予以头孢他啶抗感染治疗后,患者体温及痰量一度得到控制,但后期再次反复发热,痰量增多。治疗方案先后使用注射用哌拉西林他唑巴坦联合依替米星、注射用美罗培南,感染情况仍未明显好转。近期患者连续两次痰培养显示耐碳青霉烯类鲍曼不动杆菌(亚胺培南MIC=8 μg/L,哌拉西林他唑巴坦MIC>128 μg/L)。遂选用头把他啶阿维巴坦静脉抗感染,1天后患者体温降至正常,2天后炎症指标无明显升高。
分析
患者急性起病,初步诊断为社区获得性肺炎,细菌感染。患者老年男性,有基础疾病,在革兰阳性球菌感染可能外,革兰阴性杆菌感染可能性也存在。
初步抗感染治疗选择超广谱的第三代头孢菌素头孢他啶。感染情况没有明显控制,遂予以哌拉西林他唑巴坦与依替米星的联合治疗方案。仍未好转,考虑产β-内酰胺酶的革兰阴性杆菌感染可能性大。黏菌素类、替加环素治疗可能有效,但需监测指标较多且药物治疗窗较窄,所以治疗方案升级为抗菌活性更强的碳青霉烯类抗菌药美罗培南。
近期回报的微生物检验报告显示感染菌为耐碳青霉烯类鲍曼不动杆菌(CRAB)。鲍曼不动杆菌是临床最难治的致病菌之一,其分离率、感染率、耐药性均呈上升趋势。一旦鲍曼不动杆菌表现出碳青霉烯类的耐药性,通常会对多数其他预期对鲍曼不动杆菌有活性的抗生素发生耐药,导致剩下的治疗选择很少。目前没有明确针对CRAB感染的标准抗生素方案来评估各种治疗方案的有效性。
结合上述治疗方案考虑其耐药机制是产A类丝氨酸碳青霉烯酶的KPC酶或D类丝氨酸碳青霉烯酶的OXA酶,遂选用对两类耐药机制均有活性的头孢他啶阿维巴坦。最终患者的感染得到控制,病情得以逐渐好转。
预防
细菌耐药机制种类繁多,包括膜屏障机制,使抗菌药物无法进入细菌体内;产生灭活酶,灭活进入体内的抗菌药物;作用靶位突变,使抗菌药物无法与作用位点结合;形成生物膜,阻断细菌与抗菌药物接触;主动休眠,产生芽孢休眠体,抵御抗菌药物入侵体内。本案例中鲍曼不动杆菌产生了β-内酰胺酶,从而水解抗菌药物中的β-内酰胺环,使抗菌药物失去效力。
下面我们来了解一下常见β内酰胺酶及其抑制剂特点。
β-内酰胺酶是由细菌产生的,能水解β-内酰胺类抗生素的一大类酶。β-内酰胺酶种类繁多,有多种分类方法,最主要的分类方法有两种:一是根据β-内酰胺酶的底物、生化特性及是否被酶抑制剂所抑制的功能分类法(Bush分类法),其将β-内酰胺酶分为青霉素酶、广谱酶、超广谱β-内酰胺酶(ESBLs)、头孢菌素酶(AmpC酶)和碳青霉烯酶等;二是根据β-内酰胺酶末端的氨基酸序列特征的分子生物学分类法(Ambler分类法),将β-内酰胺酶分为丝氨酸酶(包括A类、C类酶和D类酶)及金属酶(B类酶)。以下对临床常见的三类β-内酰胺酶进行简单介绍。
1、ESBLs
是由质粒介导的能水解青霉素类、头孢菌素及单环酰胺类等β-内酰胺类抗生素的β-内酰胺酶。
其对碳青霉烯类和头霉素类水解能力弱。
主要由肠杆菌科细菌产生,以肺炎克雷伯菌、大肠埃希菌、变形杆菌最为常见。
根据编码基因的同源性,ESBLs可分为TEM型、SHV型、CTX-M型、OXA型和其他型共5大类型。
2、AmpC酶
属C类酶,通常由染色体介导,可以被β-内酰胺类抗生素诱导。部分由质粒介导,常呈持续高水平表达。
对第一、二、三代头孢菌素水解能力强,但对碳青霉烯类抗生素和第四代头孢菌素的水解能力弱。
主要存在于肠杆菌属、柠檬酸杆菌属、普鲁菲登菌属、黏质沙雷菌属和摩根菌属等细菌,非发酵菌中主要见于铜绿假单胞菌。
质粒介导的β-内酰胺酶可分为CMY-2组、CMY-1组、MIR-1/ACT-1组、DHA-1组和ACC-1组等。
3、碳青霉烯酶
能水解碳青霉烯类抗生素的一大类β-内酰胺酶,分别属于Ambler分子分类中的A类、B类和D类酶。
A类、D类为丝氨酸酶。
B类为金属酶,以锌离子为活性中心。
A类碳青霉烯酶
由染色体介导,也可由质粒介导。前者包括SME、NMC和IMI酶等,后者包括KPC和GES酶等。
KPC酶是近年来肠杆菌科细菌尤其是肺炎克雷伯菌对包括碳青霉烯类抗生素在内的几乎所有β内酰胺类抗生素耐药的最主要机制,我国最常见的是KPC-2,其对头孢吡肟和头孢他啶的水解能力相对较弱。
可被新型酶抑制剂阿维巴坦、雷利巴坦和法硼巴坦抑制,部分被克拉维酸所抑制。
B类碳青霉烯酶(金属酶)
能灭活青霉素类、头孢菌素类、碳青霉烯类抗生素,但对氨曲南水解活性弱,不能被β内酰胺酶抑制剂所抑制,可被EDTA或巯基类化合物抑制。
常见于铜绿假单胞菌、不动杆菌属细菌和肠杆菌科细菌,包括IMP、VIM、GIM、SPM、SIM、NDM酶等。
D类碳青霉烯酶(OXA酶)
对苯唑西林水解活性强,主要见于不动杆菌属细菌。
包括OXA-23、OXA-24/OXA-40、OXA-48、OXA-58 和OXA-51 酶等。
目前临床应用的酶抑制剂对其没有很好的抑制作用(除OXA-48可被阿维巴坦抑制外)。
不同OXA酶对碳青霉烯类抗生素水解活性不相同,β-内酰胺酶抑制剂的抑酶活性也不同。
β-内酰胺酶抑制剂(BLI)通常都是与β-内酰胺类抗生素结合发挥作用,通过与β-内酰胺酶的活性位点相结合,有效抑制β内酰胺酶,使得抗生素中的内酰胺环不被水解,从而保持抗生素结构的完整性而产生灭菌效果。目前临床常见的BLI主要有克拉维酸、舒巴坦、他唑巴坦和阿维巴坦。
克拉维酸(clavulanic acid)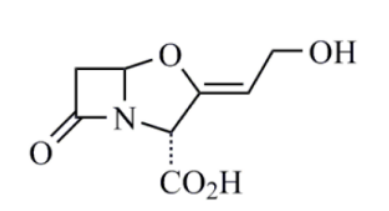
克拉维酸对质粒介导的 β-内酰胺酶具有很强的抑制活性,但对染色体介导的 β-内酰胺酶无效。
舒巴坦(sulbactam)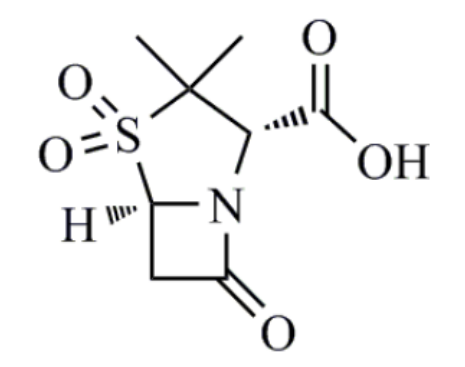
舒巴坦稳定,半衰期长,除了质粒 β-内酰胺酶外,对染色体产生的诱导酶也有一定的抑制作用。与其他BLI不同的是,舒巴坦自有一定的抗菌活性,对淋病奈瑟菌、脑膜炎奈瑟菌和不动杆菌有较强的抗菌活性。
他唑巴坦(tazobactam)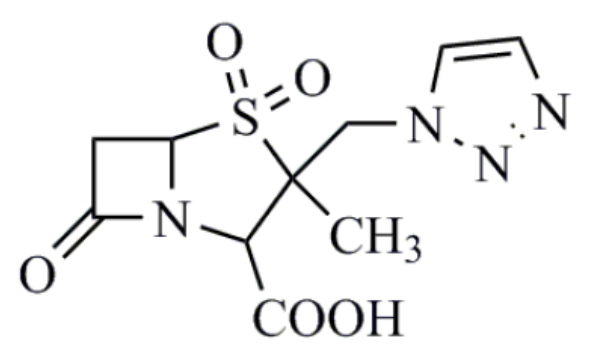
他唑巴坦抑酶谱更广、组织穿透性强、分布广。它对染色体、质粒介导的及多种超广谱酶、青霉素酶均有抑制作用。诱导细菌产酶方面,他唑巴坦明显低于克拉维酸及舒巴坦,对细菌耐药诱导能力极低,产生β-内酰胺酶的变异菌株频率亦低。
阿维巴坦(avibactam)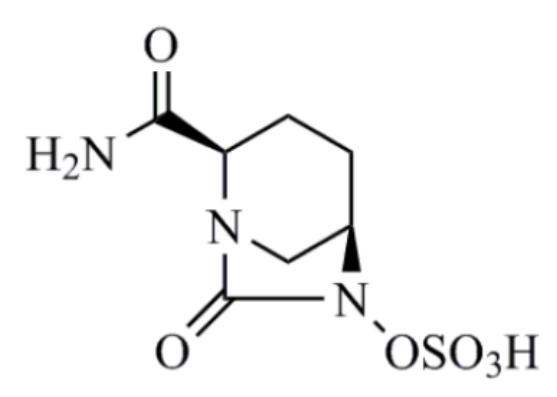
阿维巴坦不属于β-内酰胺类,与经典β-内酰胺酶抑制的作用机制不同,阿维巴坦自身结构不可恢复,因而具有长效的抑酶作用,阿维巴坦本身没有明显的抗菌活性,能抑制A类(包括ESBLs和KPC),C类和部分D类β-内酰胺酶。
不同β-内酰胺酶抑制剂特点如下:
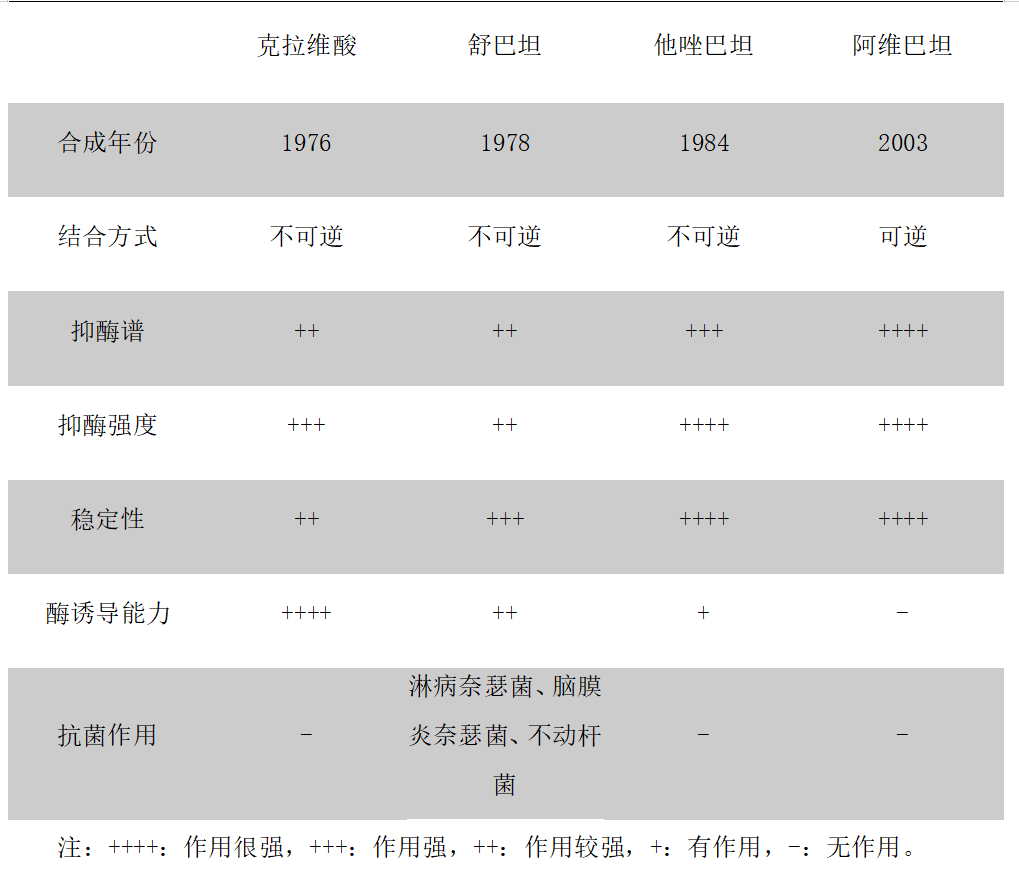
抑酶活性如下:
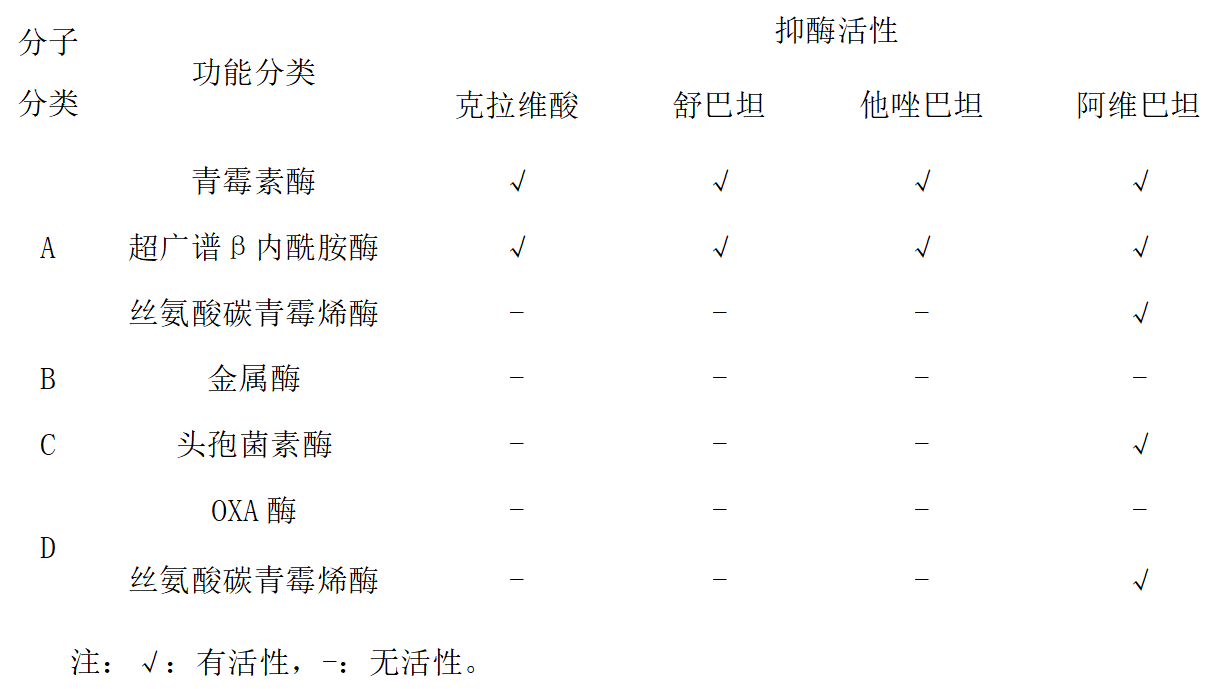
1.β-内酰胺酶按分子生物学分类法分为哪几类?
2.舒巴坦单药对哪些细菌具有一定抗菌活性?
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017