



200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过




王 钢1 王谊晴2 梁 菊1 综述 刘志红1,2 审校
DOI:10.3969/j.issn.1006-298X.2024.03.010
[基金项目]国家自然科学基金(32141004, 82300800);国家重点研发计划(2021YFC2501302);江苏省自然科学基金(BK20210150)
[作者单位]1东部战区总医院 国家肾脏疾病临床医学研究中心 (南京,210016);2浙江大学医学院
摘 要
肾脏类器官领域的快速发展,为人类肾脏发育及肾脏疾病的研究提供了重要模型支持。肾脏类器官研究已从早期建立肾单位类器官为主,发展到输尿管胚芽/集合管类器官,从而可以更全面地模拟人类肾脏,使其更好地应用于肾脏发育模拟,疾病模型及药物筛选领域。此外,单细胞测序、多组学联用、基因编辑、器官芯片等新技术的交叉融合,使得对肾脏类器官的认识更全面和深入。本文将简介肾单位类器官以及输尿管胚芽/集合管类器官诱导方法、肾脏类器官细胞及结构鉴定优化、肾脏类器官的相关应用及改进措施。
关键词 肾脏类器官 诱导方法 疾病模型
Research and application of kidney organoids
WANG Gang1, WANG Yiqing2, LIANG Ju1, LIU Zhihong1,2
1National Clinical Research Center for Kidney Diseases, Jinling Hospital, Nanjing 210016, China
2 Zhejiang University School of Medicine, Hangzhou 310058, China
ABSTRACT
The rapid development of the field of kidney organoids has provided important model support for the study of human kidney development and kidney diseases. The research on kidney organoids has evolved from early establishment of nephron organoids to ureteric bud/collecting duct organoids, which can more comprehensively simulate human kidneys and better apply them in the fields of kidney development simulation, disease modeling, and drug screening. In addition, the cross fusion of new technologies such as single-cell sequencing, multi omics combination, gene editing, and organ-chips has also enabled researchers to have a more comprehensive and in-depth understanding of kidney organoids. This article will introduce the induction methods for nephron organoids and ureteric bud/collecting duct organoids, optimization of cell and structure for kidney organoids, relevant applications and improvement measures for kidney organoids.
Key words kidney organoid differentiation protocol disease model
肾脏类器官是在体外采用胚胎干细胞(hESCs)或诱导多能干细胞(iPSC)逐步诱导分化形成的由足细胞、肾小管上皮细胞、内皮细胞和间质细胞组成的多细胞3D模型[1]。早期研究者主要以建立肾单位类器官为主,但近年来,已经有多个团队报道输尿管胚芽/集合管类器官的方法。肾脏类器官目前被广泛用于疾病建模和药物筛选,此外研究者也在积极探索肾脏类器官临床移植的可行性。本文主要介绍近年来肾脏类器官研究领域的最新进展,包括肾脏类器官诱导方法、肾脏类器官细胞及结构鉴定优化、肾脏类器官的相关应用、肾脏类器官的多组学分析及肾脏类器官研究领域的限制和挑战。
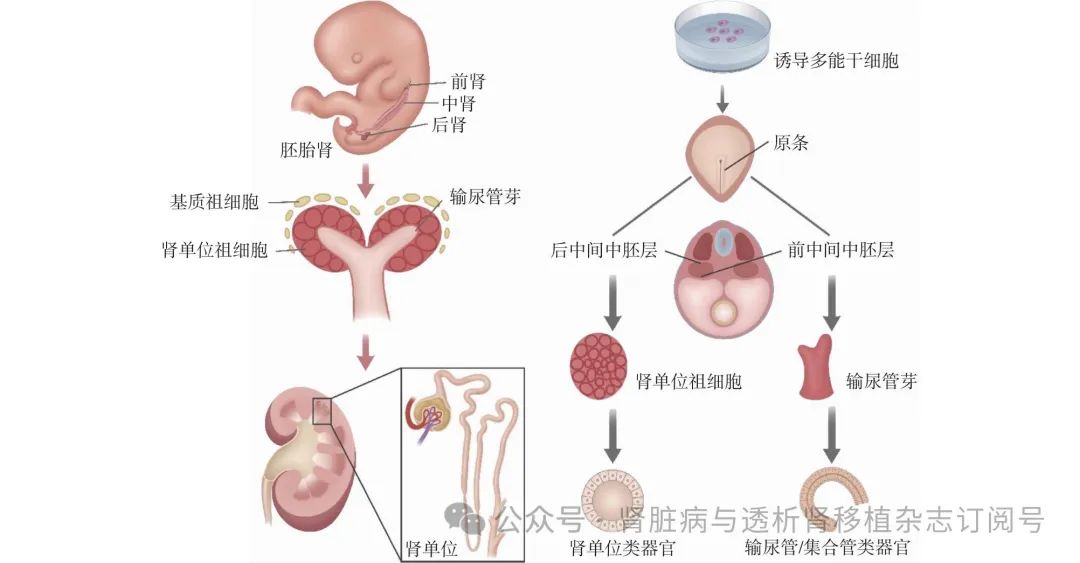
肾脏发育成熟过程 人类胚胎时期的肾脏发育可分为前肾、中肾及后肾三个阶段。前肾发生始于妊娠第22天,并于第4周末逐渐开始退化,其前肾管下端继续向尾侧延伸,成为中肾管。中肾发生于妊娠第24天,至妊娠的第3个月已大部分退化,在男性中,性腺区域的中肾小管分化形成输精小管。后肾发生始于妊娠第28天,成熟肾脏细胞类型主要由后肾中四种前体细胞发育产生:分化为肾单位祖细胞、分化为集合管和输尿管的输尿管芽、分化为肾基质/间质的基质祖细胞,以及分化为血管内皮细胞的内皮祖细胞。肾单位祖细胞和输尿管芽的分化发育在空间位置上是不同的,因此两者在中胚层的起源位置也不相同:肾单位祖细胞来源于后中间中胚层,而输尿管芽来源于前中间中胚层。肾脏类器官的诱导过程正是通过模仿这些前体细胞的发育过程而产生的,因此也就产生肾单位类器官以及集合管/输尿管类器官(图1)。
图1 肾脏发育过程及类器官的诱导流程
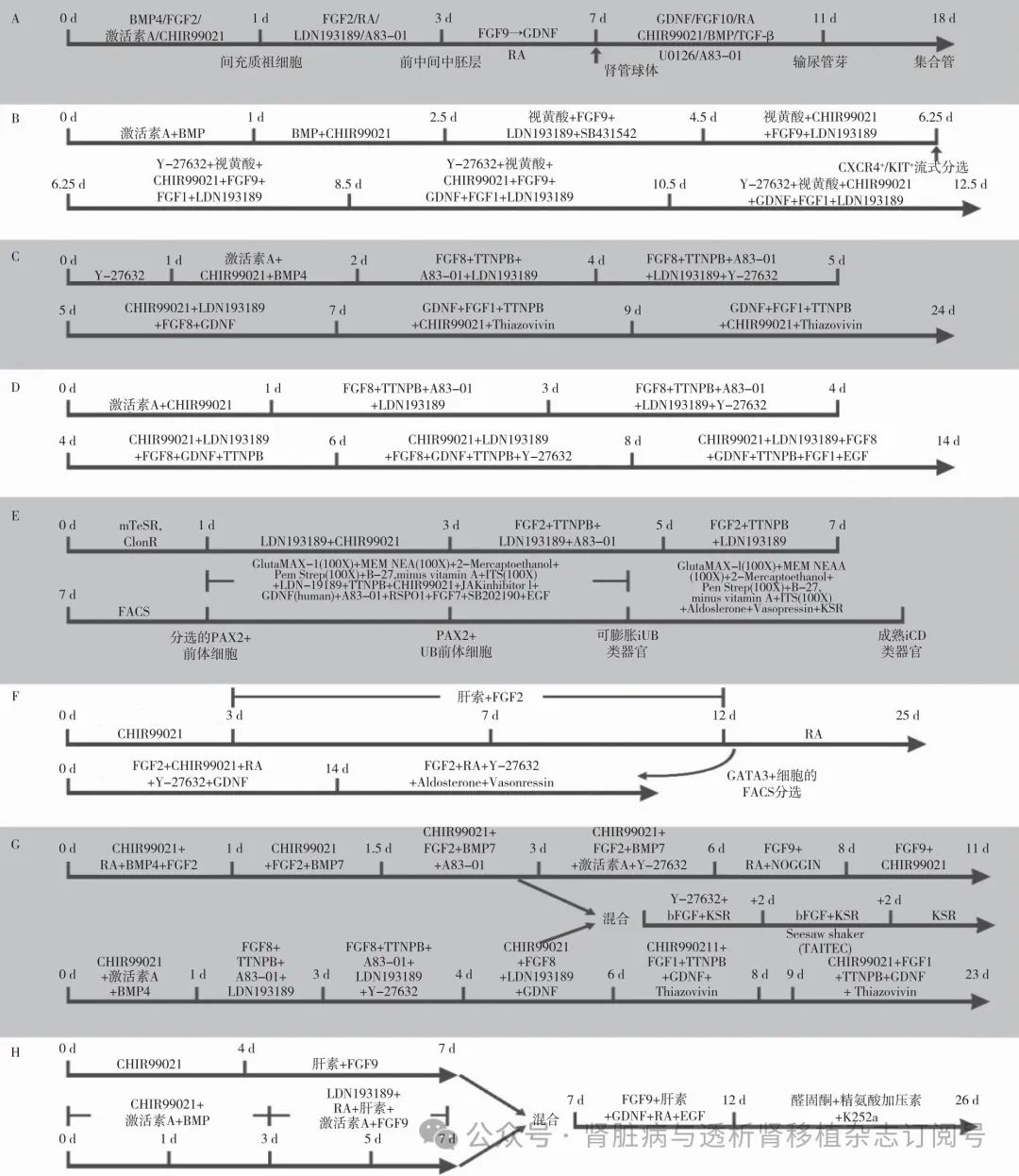
肾单位类器官诱导方法 肾单位类器官的诱导路径:iPSC-原条-后中间中胚层-肾单位祖细胞-肾单位相关细胞类型。已报道的诱导方法均基于此发育过程,在对应的分化阶段加入诱导分子。各个方法间的主要不同在于诱导分子的种类、处理时间、处理浓度及最后形成类器官的培养方式。诱导方法总结于图2[2-9],下文以Melissa H Little团队诱导方法为例进行介绍[4]。
图2 肾单位类器官诱导方法
CHIR99021:Wnt信号通路抑制剂;Noggin:BMP抑制剂;Dorsomorphin:AMPK抑制剂;BMP4:骨形态发生蛋白4;FGF:成纤维细胞生长因子;PVA:聚乙烯醇;MC:甲基纤维素;CDBLY2:培养基及诱导因子混合物;SB:转化生长因子β(TGF-β)受体激酶抑制剂;IWR-1:端锚聚合酶抑制剂;RA:维甲酸;A:Melissa H Little团队肾单位类器官悬浮诱导方法流程[2];B:Melissa H Little团队近端小管特异性肾单位类器官诱导方法流程[3];C:Melissa H Little团队肾单位类器官Transwell小室诱导方法流程[4];D:Joseph V Bonventre团队肾单位类器官诱导方法流程:2D以及3D培养模式均有介绍[5];E:Yong-Mahn Han团队肾单位类器官诱导方法流程:需与小鼠胚胎脊髓共培养[6];F:Nuria Montserrat团队肾单位类器官诱导方法流程[7];G:Yun Xia团队肾单位类器官诱导方法流程:类器官血管化水平较高[8];H:Nuria Montserrat团队肾单位类器官诱导方法流程[9]
iPSC种板之后,采用WNT通路的抑制剂CHIR99021处理3~4 d,将其诱导形成中间中胚层。然后利用成纤维细胞生长因子(FGF)9和肝素处理3 d使其进一步分化形成肾单位祖细胞。之后将肾单位祖细胞消化,并重新聚团培养于Transwell小室中5 d,延续FGF9和肝素的处理。随后撤去所有诱导因子维持培养至25 d形成类器官。该团队对以上类器官方法进行优化,建立效率更高、成本更低的悬浮培养方式[2]。在贴壁培养7 d之后,将肾单位消化成单细胞,并悬浮培养于低吸附板中,采用FGF9和肝素处理5 d之后撤去诱导因子。随后维持培养至25 d会形成小球样的肾脏类器官。
该团队还建立近端小管特异性的肾脏类器官,弥补之前肾脏类器官中近曲小管不成熟的缺点。该方法前7天诱导中间中胚层过程与之前方法一致,之后采用CDBLY2处理5 d形成肾单位祖细胞。将肾单位祖细胞消化,并重新聚团培养于Transwell小室中5 d,诱导因子为FGF9和肝素。之后撤去所有诱导因子维持培养至25 d,将会形成近曲小管特异的肾脏类器官[3]。
输尿管/集合管类器官诱导方法 考虑到输尿管芽和后肾间质分别来源于中间中胚层的前部和后部,两者的定向分化策略在诱导早期阶段有所不同。中胚层在发育过程中以头端到尾端的方式构建,其中前部中间中胚层来源于相对早期的原条。因此,原条短时间的处理会诱导分化形成中间中胚层的前部。下文以Joseph V. Bonventre团队诱导方法为例进行介绍[10],其余诱导方法总结于图3[11-17]。
图3 输尿管/集合管类器官诱导方法
BMP:骨形态发生蛋白;TGF-β:转化生长因子β;FGF:成纤维细胞生长因子;RA:维甲酸;GDNF:胶质细胞源性神经营养因子;Thiazovivin:Rock抑制剂;TTNPB:芳香维甲酸;LDN-19189:BMP4抑制剂; SB202190:选择性p38MAPK抑制剂;EGF:表皮生长因子;Aldoslerone:醛固酮;Vasopressin:血管升压素;2-Mercaptoethanol:2-巯基乙醇;A83-01:TGF-β 抑制剂;RSPO1:WNT通路激活剂;A:Joseph V. Bonventre团队输尿管/集合管类器官诱导方法流程[10];B:Ryuichi Nishinakamura团队输尿管/集合管类器官诱导方法流程[11];C:Kenji Osafune团队输尿管/集合管类器官诱导方法流程[12];D:Kenji Osafune团队输尿管/集合管类器官诱导方法流程[13];E:Zhongwei Li团队输尿管/集合管类器官诱导方法流程[14];F:Melissa H. Little团队输尿管/集合管类器官诱导方法流程[15];G:Kenji Osafune团队输尿管/集合管类器官诱导方法流程[16];H:Benjamin D. Humphreys团队输尿管/集合管类器官诱导方法流程[17]
iPSCs种板后,利用WNT、FGF、骨形态发生蛋白(BMP)和转化生长因子β(TGF-β)信号通路的激活剂处理1 d,诱导间充质祖细胞。然后,通过激活视黄酸和FGF途径,抑制BMP和TGF-β因子信号通路,使祖细胞立即分化成前部中间中胚层。在分化的第3天使用低附着板培养中间中胚层细胞形成3D小球。之后4 d,球体会表达肾管的标志物PAX2、GATA3以及 ALDH1A24。输尿管树的形态发生是由输尿管胚芽尖端的分叉分支驱动的,在体内,分叉分支由后肾间质邻近的帽状间充质细胞引导。促进分支的关键旁分泌因子包括胶质细胞源性神经营养因子(GDNF)和FGF信号。在培养的第7天将肾管球体嵌入细胞外基质,并利用多种诱导因子[如GDNF、FGF10、CHIR99021、维甲酸(RA)、U0126及A83-01]诱导分化。1周后,经历几轮分支形成输尿管芽的经典表型。
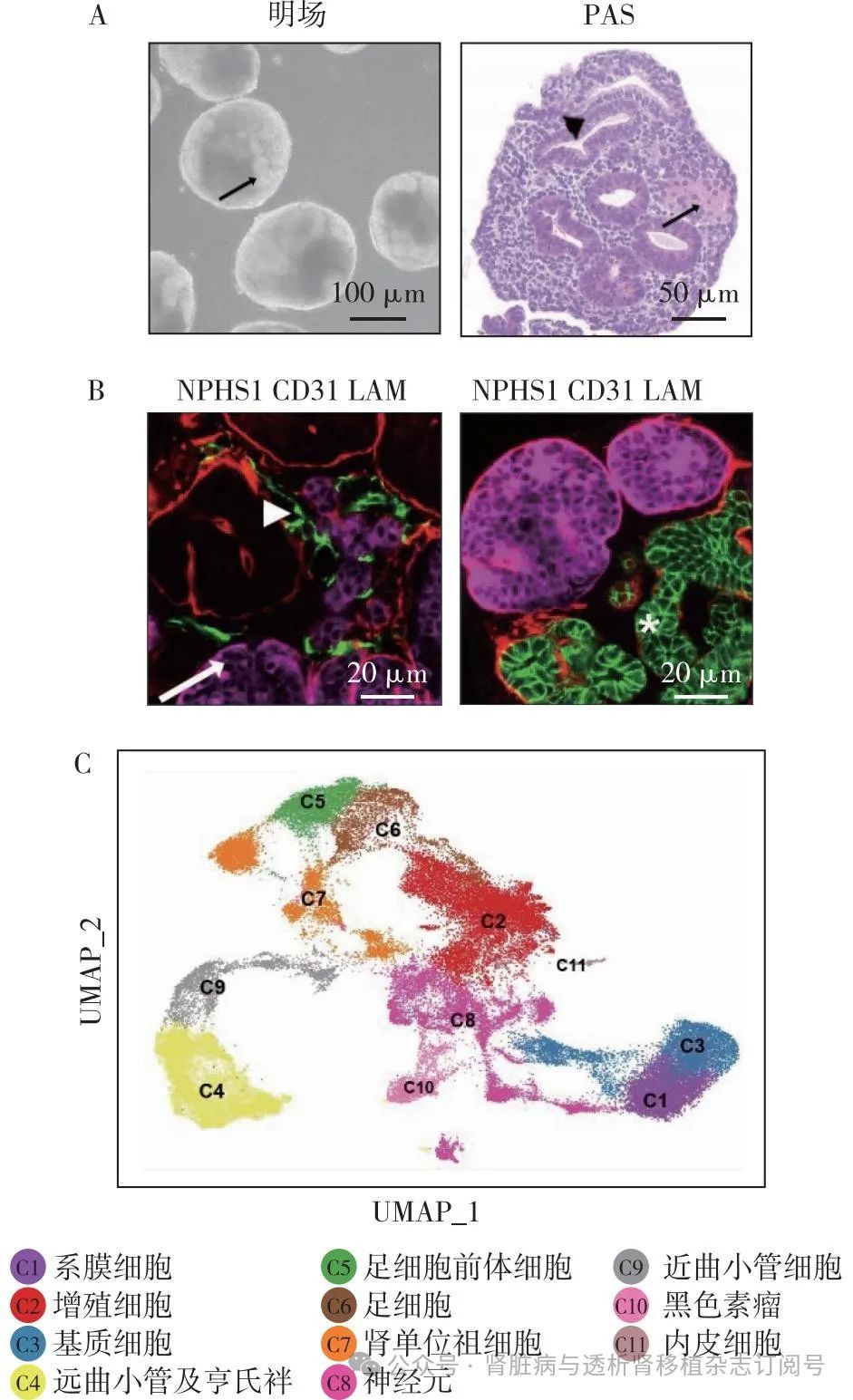
肾脏类器官的组织结构 肾脏类器官中存在肾小球、肾小管结构以及部分的基质细胞。但是由于没有血管的形成,肾小球是由足细胞团构成,并不存在毛细血管袢以及肾小球滤过屏障结构。超微结构显示,其存在明显的足细胞团、足细胞足突(次级足突纵横交错)以及裂孔隔膜。肾小管形成刷状缘结构,并且具有初步的重吸收功能[8]。
肾脏类器官的细胞类型 本研究团队的单细胞转录组学(scRNA-seq)以及免疫荧光结果显示,不同诱导方法产生的肾脏类器官中均包含肾单位祖细胞、内皮细胞、足细胞、肾小管细胞以及基质细胞(图4)[18]。除以上肾脏固有细胞类型外,还发现类器官中存在一定数量的脱靶细胞如胶质细胞、神经细胞和肌肉祖细胞。有研究整合肾脏类器官与孕期17周的胎儿肾脏单细胞数据,分析表明类器官中基质细胞、肾单位相关细胞和内皮细胞与胎儿肾脏细胞基本相似[19]。
NPHS1:足细胞标志物;CD31:内皮细胞标志物;ECAD:肾小管细胞标志物;LAM:肾小球基膜标志物;A:肾脏类器官明场及PAS染色;B:肾脏类器官免疫荧光染色;C:肾脏类器官单细胞测序
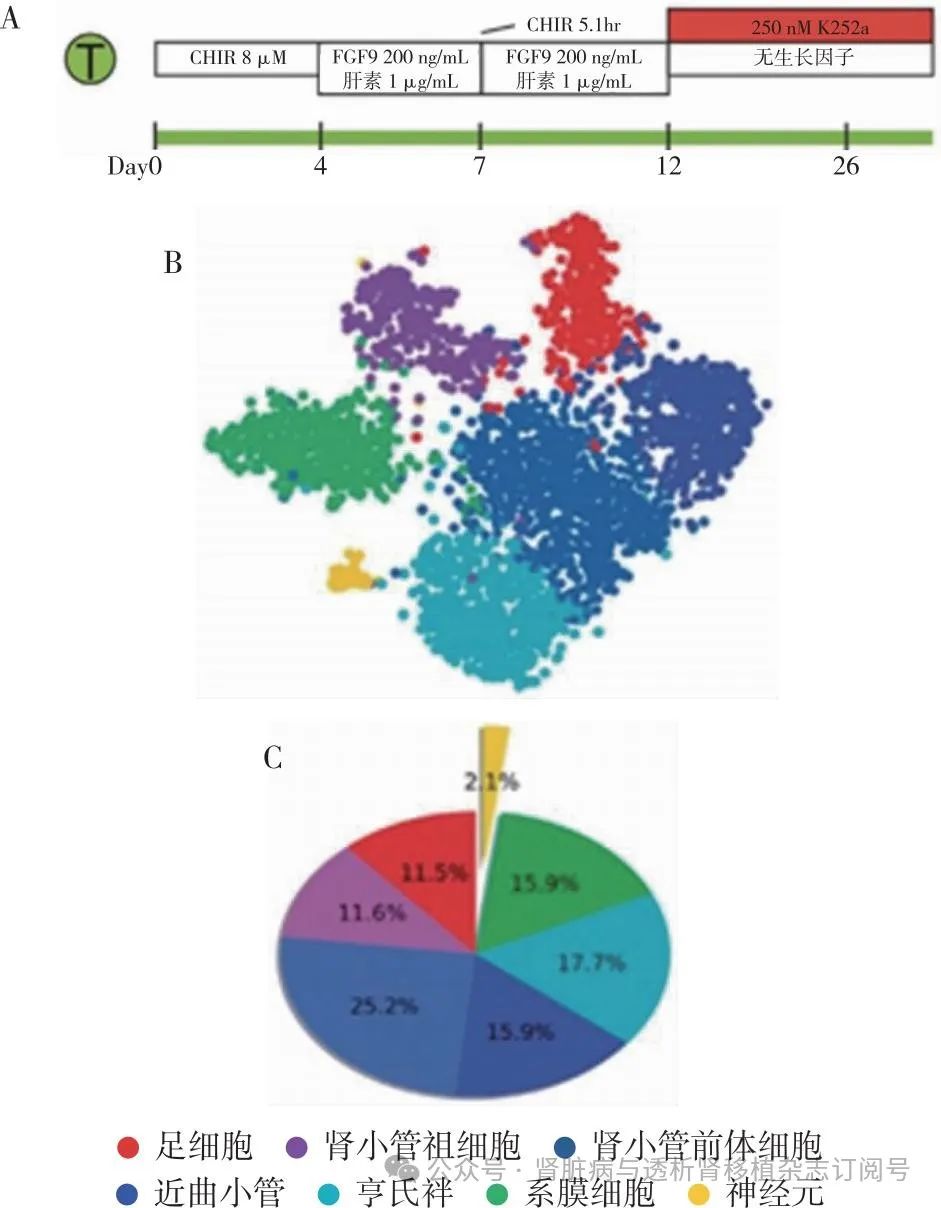
肾脏类器官的优化 肾脏类器官的不足之处主要包括脱靶细胞和成熟较低两方面。针对以上问题,很多团队对类器官的诱导进行改进。Wu等[20]研究利用scRNA-seq拟时序分析发现,类器官在诱导的中期,足细胞发育路径上出现脱靶分化效应产生神经细胞。同时,该研究团队发现BDNF及其同源受体NTRK2在类器官分化过程中的神经元谱系中表达。抑制该途径可在不影响类器官分化的情况下减少90%的神经元细胞,从而改善类器官的形成(图5)。
图5 抑制神经细胞产生的类器官诱导方法[19]
FGF:成纤维细胞生长因子;A:肾脏类器官诱导流程;B:肾脏类器官单细胞转录组t-SNE分群图;C:神经细胞比例下降至2.1%
类器官另一个不足主要表现在成熟度低及没有血管化的肾小球。scRNA-seq测序结果显示,体外培养的肾脏类器官中存在一定比例的肾单位祖细胞。并且足细胞还表达未成熟的标志基因OLFM3[19]。此外,组织结构显示,类器官中并没有成熟的肾小球滤过屏障。本团队及其他研究者将体外培养的肾脏类器官植入到免疫缺陷小鼠的肾膜下,使其与小鼠肾脏形成血液循环。该操作可使类器官进一步成熟,形成血管化的肾小球和成熟的滤过屏障。尾静脉注射葡聚糖实验可以证明类器官与小鼠肾脏形成了血液循环,并且可以通过荧光强度的变化计算类器官肾小球的滤过率[21]。除体内移植之外,也有研究团队在体外培养阶段探索增加肾脏类器官血管化及成熟度的可行性。将肾脏类器官移植在鸡胚绒毛尿囊膜上进行培养,可促进细胞的成熟及类器官的血管化[9]。根据鸡胚绒毛尿囊膜的硬度建立的水凝胶同样可以起到促进类器官成熟的作用。此外,将类器官种植于微流控的器官芯片上,利用培养基流动产生的切力可有效地促进类器官的血管化及成熟程度[22]。有研究团队,将猪的肾脏进行去细胞处理,利用剩下的细胞基质结构制作成基质胶。采用该基质胶培养肾脏类器官可显著促进类器官的血管化水平[23]。
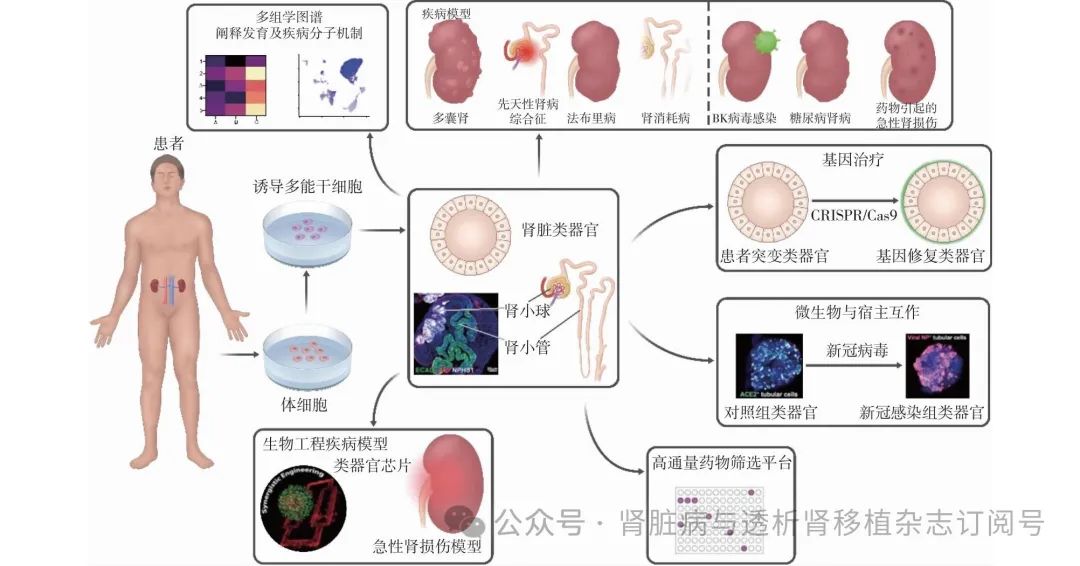
目前,肾脏类器官主要应用于疾病模型、药物肾毒性及有效性筛选(图6)。传统二维培养的肾脏细胞主要来源于永生化的细胞系或者原代细胞。但是这两者都存在着明显不足。与永生化足细胞系转录谱相比,类器官中的足细胞与成人足细胞相似程度更大。另外,人原代肾脏细胞不仅来源比较少,并且长时间的培养会出现去分化的状态。因此,肾脏类器官在肾脏领域研究应用上具有很大优势。
图6 肾脏类器官的应用
遗传性肾脏疾病模型
多囊肾 常染色体显性遗传多囊肾病(ADPKD)结构上的改变是较容易被识别的表型,有研究将PKD1或PKD2的截短突变引入iPSC,并将其分化为肾脏类器官。PKD1/2敲除的类器官自发地形成囊肿。也有研究利用ADPKD患者来源的iPSC诱导形成肾脏类器官,同样能够观察到囊肿的形成[24]。此外,有研究利用ADPKD患者iPSC诱导产生输尿管/集合管类器官,同样能够在输尿管胚芽茎部区域观察到囊肿形成[17]。
纤毛类疾病 有研究利用携带IFT140突变的纤毛病患者来源的iPSC诱导形成肾脏类器官。该类器官小管中的初级纤毛呈缩短的棒状,基因编辑修正该突变之后可以逆转这种表型。进一步将患者与修正类器官进行差异基因表达分析表明,该突变主要影响上皮小管细胞中细胞连接和动力蛋白运动组装相关等信号通路[25]。
法布里病 有研究建立α半乳糖苷酶(GLA)基因敲除的iPSC,并将其诱导形成肾脏类器官。GLA突变的肾脏类器官中足细胞和小管细胞出现三糖基神经酰胺(Gb3)的积累。超微结构分析显示,足细胞和小管细胞中有大量的电子致密颗粒沉积物和电子致密片层脂质样沉积物。GLA敲除及突变肾脏类器官的氧化应激水平升高,并伴细胞凋亡,模拟该病的疾病表型[26]。
Alport综合征 本研究团队建立了来源于Alport综合征患者的肾脏类器官疾病模型。我们将携带COL4A5突变的男性X连锁遗传Alport综合征(XLAS)患者外周血单个核细胞(PBMC)重编程获得iPSC,并定向诱导分化形成肾脏类器官。COL4A5突变不影响肾脏类器官的诱导分化,AS患者类器官具备足细胞、肾小管细胞等关键细胞类型,并在植入体内后形成成熟的肾小球基膜(GBM)。利用免疫荧光从蛋白层面观察,类器官GBM中存在α5链缺失,证明Ⅳ型胶原三聚体无法完成从col(Ⅳ)α1α2α1到col(Ⅳ)α3α4α5的转变;然而,scRNA-seq结果提示,从体外培养阶段到体内植入阶段,AS类器官和WT类器官一样,存在COL4A3、COL4A4及COL4A5表达上调,COL4A1及COL4A2表达下调。因此,提示GBM中col(Ⅳ)α3α4α5的缺失可能受转录下游事件引的调控(未发表资料)。
遗传性肾病综合征 滤过屏障是肾小球发挥滤过作用的关键结构,其中由足细胞足突间形成的裂孔隔膜发挥关键作用。NPHS1是足细胞外形成裂孔隔膜关键跨膜蛋白之一。有研究建立了携带NPHS1基因突变的iPSC, 并诱导形成肾脏类器官。免疫荧光及超微结构结果显示,患者类器官的裂孔隔膜消失。同时,利用CRISPR-cas9技术将NPHS1突变修正模型之后,该疾病表型也得到修复[27]。
遗传性肾小管间质疾病 有研究建立了携带MUC1突变患者的肾脏类器官疾病模型。利用该患者肾脏类器官揭示了MUC1基因突变通过激活未折叠蛋白反应(UPR)通路导致肾小管细胞损伤。结合高内涵筛选技术发现,BRD4780小分子可剂量依赖地减少MUC1错误折叠蛋白的蓄积,进而缓解肾小管损伤[28]。
其他 由于器官发育不成熟、缺乏血液灌注以及与免疫系统的互助,肾脏类器官目前很少用于非遗传性疾病的建模。尽管如此,一些研究还是成功地利用肾脏类器官来模拟损伤因素与肾脏的相互作用。BK病毒感染是肾移植失败的常见原因,有研究用BK病毒感染成人肾上皮细胞衍生的小管模型。受感染的肾小管呈现增大的细胞核,并出现DNA片段[29]。糖尿病肾病是发病率很高的继发性肾脏疾病,有研究利用周期性的高糖低糖处理肾脏类器官,建立糖尿病肾病模型[30]。该模型能够模拟肾脏损伤及纤维化等疾病表型。
药物肾毒性检测 肾脏是抗肿瘤药物及其代谢产物的主要清除器官。抗肿瘤药物可直接影响肾小球、肾小管及间质等结构,导致从无症状的血清肌酐升高到需要透析的急性肾功能衰竭等不同严重程度的肾脏疾病。研究者利用患者来源的肾脏类器官作为工具,在体外检测治疗药物对于该患者是否具有肾毒性,进而为肿瘤患者提供既有效同时肾毒性小的治疗方案。前期研究者利用肾脏类器官检测抗肿瘤药物庆大霉素和顺铂的药物肾毒性,两药物处理肾脏类器官之后,类器官整体结构出现损伤,细胞活性明显下降。足细胞标志物NPHS1及MAFB表达均下调,而肾损伤标志物1和凋亡标志物表达均升高[31]。以上肾脏类器官的细胞表型及分子水平的变化,可以作为在体外评估药物肾毒性的重要指标。
治疗药物筛选 近期,美国食品药物监督管理局(FDA)不再要求在药物临床试验前进行动物试验,并积极推动类器官以及器官芯片在药物临床试验前的应用。囊性纤维化跨膜传导调节因子(CFTR)抑制剂成功阻断ADPKD和常染色体隐性遗传多囊肾病(ARPKD)患者肾类器官中的囊肿形成。此外,毒胡萝卜素(Thapsigargin)分子通过抑制内质网Ca2+ATP酶,抑制ARPKD类器官中的囊肿形成[8]。除了验证对囊肿发生具有已知影响的化合物外,多囊肾类器官还能够筛选对囊肿形成具有未知影响的化合物。Blebbistatin是一种非肌肉肌球蛋白II抑制剂,可显著诱导多囊肾类器官中囊肿的形成[32]。近期研究显示,植入后类器官可与宿主形成血液循环,并且类器官结构更加成熟,可作为药物筛选更好的模型。经灌胃给药后,能够在移植后的类器官中检测到药物的存在。同时,该研究利用硫酸苯酯诱导类器官损伤模型后,经灌胃给药选择性TRPC5抑制剂(GFB-887)小分子能够缓解移植后类器官的损伤[33]。
随着测序技术的发展,多组学分析广泛地应用到肾脏类器官的优化及相关疾病模拟的研究当中。肾脏类器官的蛋白质组学显示,类器官中可以表达肾小球及肾小管基膜相关组分:COL4A1、COL4A2、COL4A3、COL4A5及COL4A6等。该研究进一步利用scRNA-seq分析以上蛋白组分的细胞来源,明确不同细胞类型在基底膜形成过程中所发挥的作用,如COL4A3/A5两种基因只在足细胞中特定表达。
此外,有研究利用scRNA-seq和单细胞染色质可及性测序(scATAC-seq)对类器官的分化过程在表观基因组和转录组水平上进行了详细分析。该研究利用顺式可及性分析,找到HNF1B基因的增强子位置,并在近端小管细胞和类器官中证实该位置可以调控HNF1B的表达。同时,该研究还将类器官与成人肾脏进行比较,类器官细胞类型中染色质呈现更广泛开放的状态,这也符合类器官各细胞类型发育不成熟的状态[34]。
除发育分析外,有研究联合scRNA-seq和蛋白质组分析肿瘤坏死因子α(TNF-α)处理肾脏类器官之后转录组和蛋白组变化。TNF-α处理的类器官较正常组有322种差异表达的蛋白质,包括细胞因子和补体等种类。同时,这322种蛋白质的转录表达在蛋白尿性肾病临床预后较差的个体中也显著较高。之后,该研究进一步选取C3和血管细胞黏附分子1(VCAM1)两种蛋白分析发现,这两基因在人类肾小管和类器官肾单位相关细胞群中均特异性高表达。该研究表明类器官模型可以促进疾病生物标志物的发现[35]。
现阶段类器官诱导有明显的批次效应并且缺乏血供支持。不同细胞系、不同实验方法诱导形成的肾脏类器官产生的差异均小于批次效应。为解决批次效应,有团队开发了一种新的算法:Vireobulk[36],该算法可以借助样本自身的单核苷酸多态性(SNP)信息,拆分混合上样测序中的各个样本数据。该研究将对照组和疾病组类器官在iPSC状态时混合在一个孔中进行诱导,直至最终形成对照组和疾病组的嵌合肾脏类器官。最后,利用该算法拆分嵌合类器官中各个样本的数据,进而解决类器官模型批次效应。除技术层面外,肾脏类器官模型在评估肾功能方面也存在不足之处。现阶段体外培养及体内植入肾脏类器官在评价肾小球滤过及肾小管重吸收功能时,均只能通过葡聚糖实验。该实验只能定性证明肾小球的滤过及肾小管重吸收功能,与动物模型的定量指标相比差距甚远[8,21]。虽然,有研究团队利用双光子共聚焦显微镜实时地观察,荧光标记的葡聚糖在植入后类器官中荧光强度的变化,但这种测量方式受限于设备及操作误差的影响局限性太大[21]。尿液形成也是肾脏重要的生理功能,植入后肾脏类器官虽然血管化,但是缺乏完备的集合管和输尿管系统,因此不具备尿液产生的条件。
生物工程领域的发展,也推动类器官领域的发展。标准化及高通量的流程能够很好地减少实验批次之间的误差。同时,微流控的技术还能够加速类器官成熟以及血管化[22]。联合多器官类型的类器官,可以建立多系统的类器官芯片系统,为药物及疾病机制的探索提供更接近体内环境的疾病模型。
肾脏类器官已经成为模拟肾脏疾病的模型平台,但仍面临进一步完善血管化、免疫化和系统化的挑战,上述问题限制了其应用范围。随着交叉领域技术的结合,如基因编辑、单细胞水平分析、离体培养系统等,都将加速类器官领域的发展进步,在更广泛的领域为疾病发病机制研究、基因治疗、药物研发、器官功能替代提供更好的模型和路径。


来源:肾脏病与透析肾移植杂志订阅号
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017