



200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过




徐孝东 梁 菊 综述 刘志红 审校
DOI:10.3969/j.issn.1006-298X.2024.05.011
[基金项目] 江苏省肾脏病医学创新中心(CXZX202202)
[作者单位] 东部战区总医院 国家肾脏疾病临床医学研究中心(南京,210016)
摘 要 溶酶体是物质降解和回收利用的中心,错误折叠的蛋白质、病原体和受损的细胞器等均会被运输到溶酶体进行降解。此外,也是感知和协调细胞代谢、免疫反应、信号传导以及质膜修复的中心信号节点,当溶酶体功能障碍时会导致底物贮留,引发溶酶体相关疾病。已有报道,肾脏是最富含溶酶体的器官,肾脏固有细胞溶酶体结构或功能异常与多种肾脏疾病的发生发展有关。本文对溶酶体功能稳态以及稳态失衡在肾脏疾病中的作用研究进展作一综述。
关键词 溶酶体 肾脏疾病 足细胞 肾小管上皮细胞
XU Xiaodong,LIANG Ju,LIU Zhihong
National Clinical Research Center for Kidney Diseases, Jinling Hospital, Nanjing 210016, China
ABSTRACT Lysosomes play a crucial role in the degradation and recycling of cellular waste. Misfolded proteins, pathogens, and damaged organelles are transported to the lysosomes for degradation. Additionally, they serve as a central signaling hub for sensing and coordinating cellular metabolism, immune responses, signal transduction, and plasma membrane repair. Lysosomal dysfunction can lead to the accumulation of substrate within the lysosomes, giving rise to lysosomal-related diseases. Kidney cells are rich in lysosomes and previous studies have shown that abnormalities in lysosomal structure or dysfunction in kidney cells are associated with various kidney diseases. This article describes the roles of lysosomes and reviews recent studies concerning the involvement of lysosomal homeostasis disruption in kidney diseases.
Key words lysosome kidney diseases podocyte renal tubular epithelial cell
溶酶体是单层膜结构的细胞器,直径0.1~1.2 μm,哺乳动物细胞质中的数量为50~1 000个。它的组成、数量和大小都受到精细调节,能及时地对细胞内、外刺激作出反应,并根据其所处的环境而发生许多变化。近年来研究表明,溶酶体除在底物降解中的主要功能外,还在营养感知、代谢、质膜修复和免疫信号传导等方面发挥重要作用[1]。
肾脏具有滤过、重吸收和排泌等多重功能,也是溶酶体最为富集的器官。目前已发现溶酶体损伤在肾脏疾病的发生发展中起着非常重要的作用。例如,当肾小管细胞过量吸收草酸钙、白蛋白或放射性核素后,会通过溶酶体对白蛋白进行降解或通过胞吐作用将沉积的物质排出,以此减轻溶酶体膜通透性和降低细胞死亡的发生[2-4]。本文将重点介绍溶酶体功能稳态在肾脏疾病发生发展的作用。
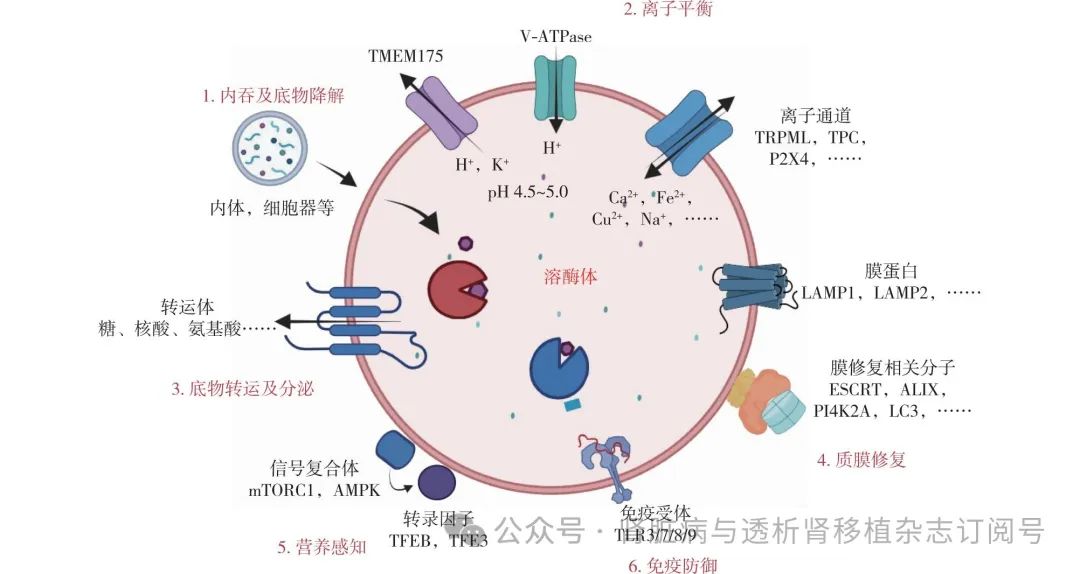
溶酶体是一种多功能细胞器,参与许多细胞重要功能,除底物降解外,还包括内吞、分泌、调节离子平衡、质膜修复、营养感知和免疫防御等重要作用。
内吞功能 被溶酶体降解的物质可分为三类:(1)通过内吞途径获取的生物小分子;(2)通过自噬途径获取的细胞内聚集体和损伤的细胞器;(3)通过吞噬途径吞噬的大颗粒物质(如细菌等)。细胞外物质的摄取和形成的质膜复合物的输入是通过动力蛋白依赖或非动力蛋白依赖机制完成的。非动力蛋白依赖性内吞作用由网格蛋白介导,有助于细胞外分子和液体的输入及质膜受体再循环。动力蛋白依赖内吞作用通过小窝蛋白介导,有助于质膜上的脂质稳态。此外,细胞膜小窝还是一些受体的驻留位点,构成了这些分子的信号平台,使得结合的分子和捕获的可溶性物质能够进入早期内体,内化的受体通过快速转运功能直接回到质膜再利用,也可通过内体循环与溶酶体融合进行降解(图1);溶酶体受损将导致生物大分子物质贮留并引发溶酶体贮积症(LSD),在细胞层面,LSD主要表现为溶酶体膨大、各类底物(如蛋白、脂类、核酸和多糖等)的积累,溶酶体降解和转运的功能下降和溶酶体再生受阻等[5]。
TMEM175:跨膜蛋白175;V-ATPase:囊泡ATP酶;TRPML:溶酶体非选择性阳离子通道;TPC:双孔离子通道蛋白;P2X4:离子通道型嘌呤受体;LAMP:溶酶体相关膜蛋白;ESCRT:内吞体运输必需分选复合物;ALIX:凋亡相关基因相互作用蛋白;PI4K2A:磷脂鞘氨醇磷酸化酶2α;LC3:蛋白轻链3;TLR:Toll样受体;TFEB:转录因子EB;TFE3:转录因子E3;mTORC1:哺乳动物雷帕霉素靶蛋白1;AMPK:单磷酸腺苷激活的蛋白激酶;1:经细胞内吞的分子被运输至溶酶体进行降解;2:离子通道维持溶酶体腔pH稳定及离子平衡;3:降解的底物经转运体转出至胞质进行再利用;4:招募相关分子用于质膜修复;5:感知细胞外环境营养物质维持细胞活性;6:降解的底物激活TLR信号轴触发天然免疫系统
离子平衡 溶酶体是H+、Ca2+、Na+、Zn2+、Cu2+、Fe2+等多种离子的储存场所[5]。比如腔室Ca2+ 浓度(0.5 mmol/L)比胞质(100 nmol/L)高5 000倍左右。溶酶体Ca2+在信号转导、转录因子调控、溶酶体重构、自噬和胞吐等方面发挥重要作用,目前已发现溶酶体Ca2+通道蛋白有溶酶体非选择性阳离子通道(TRPML)1~3、双孔离子通道蛋白(TPC)和离子通道型嘌呤受体(P2X4)[5]。研究最为清楚的TRPML1,其在机体组织细胞中广泛分布,多种物质可使其激活,比如活性氧(ROS)、溶酶体膜损伤、饥饿及磷脂酰肌醇3,5二磷酸。TRPML1基因突变可导致Ⅳ型黏脂贮积症,是一种儿童期发病的神经退行性疾病,临床主要症状为智力低下和视网膜退行性病变,是由于TRPML1功能缺陷导致溶酶体胀大、脂类物质积累以及溶酶体定位异常导致的降解功能和运输机制受损[5-6]。
正常情况下,溶酶体腔室pH值维持在4.5~5.0的超酸性环境。溶酶体酸化主要是靠溶酶体膜上的囊泡ATP酶(V-ATPase)和跨膜蛋白175(TMEM175)共同作用实现的,V-ATPase通过水解ATP 产生能量将胞质侧的H+泵入溶酶体腔内,当pH过酸时触发TMEM175通道将H+泵出以保持动态平衡[7]。
溶酶体Fe2+在腔室中的浓度为微摩尔级[5]。转铁蛋白和铁蛋白通过内吞、自噬将Fe3+带入溶酶体,Fe3+随后通过金属还原酶(STEAP3)转化为可利用的Fe2+形式,并通过溶质载体家族2成员11(SLC11A2)输出到溶酶体外发挥作用。溶酶体酸化异常会导致细胞Fe2+缺乏,而由此引起的Fe2+减少会对重要的细胞通路产生影响。
Cu2+是多种激酶和呼吸酶的辅助因子,当胞质Cu2+浓度过高时,Cu转运ATP酶β亚基(ATP7B)将胞质多余的Cu2+储存进溶酶体。Tsang等[8]研究表明,肝细胞利用溶酶体胞吐作用将Cu2+输送到胆汁中清除,以防止Cu2+浓度过高而中毒。ATP7B基因突变会导致Wilson病,是一种以Cu2+过量为特征的常染色体隐性遗传病,主要影响肝脏和大脑。
分泌功能 生理情况下存在两种类型的溶酶体,即降解型溶酶体和分泌型溶酶体,两者在形态和生化特征有共同点,如在膜表面有很多高度糖基化的蛋白,以及维持膜内较低pH 值的质子泵,膜内都具有降解蛋白质、脂类、糖的水解酶。不同之处在于分泌型溶酶体具有由一套特殊蛋白组成的致密核心颗粒,其数量和组成取决于细胞类型和细胞的活性,可受外部环境变化而释放出内容物;比如CD8 T细胞的分泌型溶酶体既可以降解大分子物质,也可在形成的免疫突触处释放内容物进行信号传递[9]。在分泌机制上,Rab蛋白家族成员27A(Rab27a)起到了核心作用,介导溶酶体所分泌的囊泡在细胞内移动和膜融合,Rab27a突变可导致多种溶酶体相关疾病的发生,如Hermansky-Pudak综合征和Griscelli等综合征[10]。
除了分泌蛋白质,溶酶体还可从细胞中分泌细胞外囊泡(包括凋亡小体、微囊泡和外泌体)[11]。其中,外泌体起源于晚期内体-溶酶体膜向内出芽过程,它不与晚期内体-溶酶体膜融合,而是在胞吐过程中以完整的囊泡形式释放到细胞外环境[11]。外泌体内含有蛋白质、脂质、miRNA等物质,其胞吐过程不仅是细胞清除废弃物的重要机制,也是细胞间通信机制。本课题组近期发现激活的CD8 T细胞可将编码的miR-186-5p以外泌体包裹的形式释放到外周血,并经循环系统被肾小管上皮细胞所吸收、触发免疫炎症反应[12]。
溶酶体膜的损伤修复 在内外刺激下,溶酶体的数量、大小和组成通过生物发生、修复或清除而不断更新。一旦溶酶体结构和功能过度和持续受损将导致细胞死亡,所以溶酶体的及时补偿和修复对于许多类型细胞的存活是非常重要的。
目前存在三种已知的溶酶体膜修复机制[1]:(1)轻度溶酶体膜损伤可以通过转运所需的内体分选复合体(ESCRT)快速、直接修复,原理是溶酶体膜结构损伤会激活ESCRT运输组分凋亡相关基因2结合蛋白X(ALIX),而ALIX又进一步招募ESCRT-Ⅲ到破裂的溶酶体膜中,ESCRT-Ⅲ复合物在膜表面形成丝状螺旋,这些螺旋通过收缩来封闭脂质双层中的漏洞。(2)当超出ESCRT修复范围时,磷脂酰肌醇-4激酶2α型(PI4K2A)会在受损的溶酶体膜上迅速积累,产生高水平的脂质信使磷脂酰肌醇-4-磷酸(PI4P),PI4P继续招募多个氧化甾醇结合蛋白家族成员,推动内质网和受损溶酶体之间的强力互作,进而激活胆固醇和磷脂酰丝氨酸转运至溶酶体,最终自噬相关基因2在磷脂酰丝氨酸的刺激下通过大规模脂质运输直接修复溶酶体漏洞。(3)当溶酶体膜损伤的持续时间和程度超过上述修复能力时会启动选择性自噬来清除永久损伤的溶酶体。这个过程涉及半乳糖凝集素的招募,它与暴露的糖基化溶酶体腔蛋白结合,并触发随后的复合物组装以促进清除。非经典的溶酶体自噬,涉及自噬微管相关蛋白轻链3(LC3)与溶酶体膜的直接结合,LC3与TRPML1相互作用促进Ca2+释放、转录因子EB(TFEB)激活和溶酶体胞吐(图1)。
营养感知 为了适应不断变化的外界环境,细胞溶酶体含有营养感知机制(图1)。当细胞营养充足时:表型上,溶酶体在大小(直径10~500 nm)、形状和位置分布等形态学上是不均匀的,每个细胞含有数百个溶酶体;在机制上,哺乳动物雷帕霉素靶蛋白1(mTORC1)会磷酸化溶酶体TFEB使其失活,而TFEB 是溶酶体生成的主要转录调节因子,其转录活性取决于其亚细胞定位,当TFEB受到抑制后会使其滞留在细胞质中,无法向核内转运行使其功能。当细胞饥饿时:表型上,溶酶体体积增大(直径500~1 200 nm),在位置分布上溶酶体会向核周区域移动,在数量上每个细胞溶酶体会降到50以下;在机制上,营养物质缺乏会激活腺苷酸活化蛋白激酶(AMPK),活化的AMPK通过抑制mTORC1使TFEB去磷酸化,去磷酸化状态的 TFEB迅速移位至细胞核并结合协调溶酶体表达和调控元件(CLEAR)后上调溶酶体相关基因表达,促进新溶酶体生成来补偿细胞溶酶体数量和质量[1]。
免疫防御 溶酶体还是细胞的免疫防御器。巨噬细胞和中性粒细胞等天然免疫细胞摄取抗原后需要形成吞噬溶酶体来降解摄入的物质,中性粒细胞也能通过释放溶酶体酶(如弹性蛋白酶)来降解细菌或内源性抗原成分(图1)。比如Toll样受体(TLR)7/8/9位于溶酶体膜中,细胞可通过TLR7/8/9识别在溶酶体内降解的病原体相关分子模式来启动天然免疫防御反应。比如CD8 T细胞分泌的miR-186-5p可激活肾小管上皮细胞内体TLR7信号轴诱发炎症因子分泌,触发肾脏局部炎症[12]。溶酶体膜损伤可释放组织蛋白酶和ROS进入细胞质,ROS一方面可诱导脂质过氧化,与游离铁积累一起导致铁下垂(自噬性细胞死亡),另一方面还能与组织蛋白酶等分子一起激活NOD样受体蛋白3(NLRP3)炎性小体,导致炎症和细胞焦亡。被细胞吞噬的病原体在溶酶体降解后还可以与MHC-Ⅱ分子结合进行抗原呈递,以触发机体的适应性免疫系统的激活。
溶酶体异常相关的肾脏疾病包括遗传与非遗传性肾脏疾病。
肾病型胱氨酸症 胱氨酸病是一种罕见的常染色体隐性遗传病,是由于CTNS基因突变导致编码的胱氨酸蛋白酶缺乏引起,使游离的胱氨酸在溶酶体内蓄积,进而导致肾功能衰竭、糖尿病、甲状腺功能减退、肌病和中枢神经系统恶化。胱氨酸病患者溶酶体功能障碍会导致肾小管上皮细胞线粒体自噬受损,破坏细胞间的紧密连接而导致上皮细胞增殖和去分化,使肾小管上皮细胞丧失重吸收能力。Rega等[13]研究发现在CTNS-/-敲除小鼠的肾小管上皮细胞中TFEB表达水平显著下调,导致溶酶体再生受损。Johnson等[14]在CTNS-/-敲除小鼠和肾病型胱氨酸症患者中发现,溶酶体Rab27a表达水平下调抑制细胞内体-溶酶体运输机制和溶酶体胞吐功能,导致胱氨酸在肾脏固有细胞中的大量蓄积;而过表达TFEB可纠正CTNS-/-小鼠肾小管上皮细胞溶酶体异常形态及数量的改变,促进溶酶体胞吐减少胱氨酸的蓄积,使超负荷的溶酶体恢复自身稳态。
法伯疾病 法伯疾病是由编码酸性神经酰胺酶基因(ASAH1)突变引起,呈常染色体隐性遗传。ASAH1基因突变导致神经酰胺在溶酶体内广泛积聚,影响多种组织器官。临床表现为声音嘶哑、皮下结节、关节挛缩畸形及中枢神经系统退行性病变。该病罕见,目前国内仅有2例散发病例报道,但均未提及肾脏是否受累[15-16]。Samuelsson等[17]报道了1例法伯疾病患者肾脏中大量神经酰胺酶积聚;最近发表的一项研究表明,将小鼠足细胞溶酶体酸性神经酰胺酶α亚基特异性敲除(Asah1fl/fl/PodoCre)后出现了严重蛋白尿,电镜下观察到明显足细胞足突融合,通过液相色谱-串联质谱法检测到小鼠肾小球中存在大量神经酰胺积聚[18]。
多囊性肾病 常染色体显性遗传性多囊肾病是终末期肾病最常见的单基因疾病,目前尚无成熟的治疗方法。在来源于内部和外部环境的刺激下,瞬时受体电位(TRP)通道能够跨膜导入阳离子,介导疼痛、温度、机械力等多种重要感知功能。多囊型TRPP是TRP家族中的重要成员,其中TRPP2(PKD2)与PKD1可发生突变并导致流体感知功能及钙信号的紊乱,进而引发多囊肾病变[19]。比如研究证实, TRPP2错义变体D511V大大降低了TRPP 2蛋白的稳定性,但溶酶体小分子抑制剂氯喹可显著抵抗TRPP2 D511V的溶酶体降解途径进而稳定TRPP2蛋白表达水平和表型上的缓解[20]。
系统性红斑狼疮(SLE) 目前在SLE等多种自身免疫性疾病中发现免疫细胞溶酶体存在功能障碍。比如在MRL/lpr小鼠脾脏中发现B细胞溶酶体体积扩大、溶酶体膜蛋白(LAMP2A,LAMP1)表达量及pH显著升高,其底物降解能力明显降低[21]。此外,因免疫细胞自噬水平缺陷引起的Th17/调节性T细胞比例失衡也是促进SLE发生发展的重要因素,在分子机制层面,MRL/lpr小鼠组织溶酶体蛋白酶K缺乏是导致狼疮肾炎的主要病理表现[22]。在SLE患者的外周血中也发现了溶酶体热休克蛋白(HSP)的自身抗体,但是否具有致病作用尚不清楚[23]。
糖尿病肾脏疾病(DKD) 众所周知,晚期糖基化终产物(AGE)是驱动DKD发生和进展的主要因素。在肾小管细胞中,暴露于微环境中的AGE被小管上皮细胞内吞触发溶酶体膜通透化,引起溶酶体酸化障碍和组织蛋白酶活性降低;增加AGE暴露时间还可进一步导致自噬底物蛋白p62的大量积聚,导致小管细胞自噬-溶酶体通路受阻而进一步加重肾损伤程度[24]。此外,DKD患者体内AGE生成过量或降解受阻还能进一步诱导细胞外基质扩张、上皮间质转分化和触发肾脏炎症等反应,使肾组织损伤进一步恶化。比如,在自噬缺陷的DKD小鼠模型中观察到AGE在肾脏血管和固有细胞中大量积聚,并伴肾脏血管中膜增厚、肾纤维化和肾脏炎症的发生[25]。体外实验中,高糖刺激肾小管上皮细胞还会导致mTOR过度活化进而使TFEB无法进行核转位,造成新溶酶体生成不足而导致数量耗竭,使自噬无法通过溶酶体途径完成[25]。
另一项研究通过整合全基因组关联研究(GWAS)和表达数量性状基因座(eQTL)数据显示编码β甘露糖苷酶基因(MANBA)是进展为慢性肾脏病的风险基因,主要分布肾小管上皮细胞的溶酶体中。为探索MANBA生理功能,通过MANBA基因敲除小鼠发现,MANBA可通过介导溶酶体结构和功能紊乱导致肾小管上皮细胞内吞功能和自噬活性降低,进而加重药物诱导的肾小管上皮细胞损伤[26]。
局灶节段性肾小球硬化(FSGS) 肾小管占肾实质90%以上,而肾小管的重吸收功能离不开溶酶体,溶酶体损伤是肾小管上皮细胞重吸收能力缺陷的基础。例如,草酸结晶被肾小管细胞吞噬后会触发溶酶体保护性反应,包括溶酶体膜的LC3脂化、TRPML1激活和TFEB核转位,通过溶酶体胞吐作用促进溶酶体再生和结晶物质的清除[3]。FSGS以大量蛋白尿起病,过量白蛋白尿会诱发氧化应激导致肾小管上皮细胞溶酶体发生膜通透化,组织蛋白酶泄漏或活性降低无法及时降解白蛋白,进而损伤小管上皮细胞。比如,将小鼠ATG5敲除会导致小管上皮细胞p62蛋白堆积、线粒体DNA损伤及功能障碍[27];也有观点认为,肾小管上皮细胞对白蛋白重吸收功能需要内体与溶酶体融合以进一步完成蛋白降解和营养物质回收,这一过程需要Ca2+通道协助,溶酶体Ca2+通道功能障碍可阻碍内体与溶酶体的融合,可能是导致肾小管蛋白尿形成的一种机制[28]。此外,化学损伤、重金属及核辐射同样可对肾小管上皮细胞溶酶体造成严重损伤。比如庆大霉素被肾小管上皮细胞吸收后会降低组织蛋白酶活性损伤溶酶体功能,同时激活caspase信号通路途径触发细胞凋亡[29];重金属铅、镉中毒会造成肾小管上皮细胞自噬受阻,导致溶酶体膜通透化继而引起细胞死亡[30];在小鼠接受急、慢性铀放射性核素污染后,大量铀会靶向肾脏并在肾小管细胞中以磷酸盐铀结晶形式累积,导致细胞溶酶体膜透化和细胞坏死,而激活TRPML1-TFEB途径不仅会提高溶酶体胞吐作用减少铀元素在细胞中的累积,还能促进溶酶体再生和质膜修复过程[4]。因此增强溶酶体功能、降低溶酶体膜通透性及组织蛋白酶泄漏有利于减轻肾脏炎症和延缓肾脏病的进展。
肾小管间质损伤 纤维化是肾脏疾病不断恶化的主要原因之一。近期研究发现在肾脏纤维化的患者及小鼠模型中观察到溶酶体内铁的大量累积,并促进ROS和衰老相关分泌表型的产生,通过磁共振成像检测铁浓度可评估肾纤维化进程[31]。肾小管细胞中的自噬对衰老和肥胖相关的肾脏疾病起着重要的保护作用,随着疾病的进展,自噬过程往往因溶酶体过载而停滞,无法进一步激活,对肾脏健康构成威胁。值得注意的是,在衰老和肥胖状态下,当自噬功能受损时,肾小管细胞会释放成纤维细胞生长因子(FGF)21,FGF-21在改善自噬停滞及促进线粒体生物合成方面显示出显著作用;而敲除FGF-21会导致老年和肥胖小鼠出现肾小管损伤,并伴随溶酶体肿胀和底物(如SQSTM 1/p62)的积累[32]。
靶向溶酶体而非上游自噬途径可能对疾病治疗更具优势,可避免具有潜在毒性作用的自噬体的积累。近期DKD的研究中,二甲双胍和白藜芦醇等AMPK激活剂作为修复自噬活性的靶点药物受到了广泛关注,AMPK激活剂通过激活自噬过程,有助于缓解DKD的病程进展[33]。此外,TFEB作为自噬与溶酶体的主要调控因子,在DKD和重金属中毒的患者发病过程中表达及活化减少,且存在入核障碍。因此,TFEB被认为是具有潜力的新治疗靶点,通过调控其表达与功能,有望为这类患者的治疗提供新的策略[25]。而在多能干细胞和类器官诱导的胱氨酸病模型中,将联合mTORC 1抑制剂(依维莫司)与半胱胺能够显著改善溶酶体结构和功能障碍,降低细胞凋亡水平[34]。另外,活性肽段P140在改善SLE患者临床表现和提高MRL/lpr小鼠存活率方面展现出了良好的前景;其作用机制涉及到升高溶酶体pH值,降低蛋白酶降解活性,以及与分子伴侣HSPA8的相互作用来降低MHC-Ⅱ分子的过度表达,从而减少自身抗原肽向T细胞的呈递[35]。而靶向调控FGF-21的停滞及自噬过程,有望开发出针对慢性肾病的新型治疗策略。这将为肾脏疾病的防治提供新的思路和方法[32]。综上所述,目前对靶向溶酶体途径的治疗研究正不断深入,新型靶点药物如TFEB激活剂、mTORC1抑制剂及活性肽段等,为改善肾脏病提供了新的可能性。
溶酶体是维持细胞稳态,代谢调节和感知环境改变的重要细胞器之一,其功能异常参与多种疾病的发生和发展,包括自身免疫疾病,代谢性疾病和肾脏疾病。进一步加强相关研究,有望为认识疾病和寻找更有效的靶向治疗提供参考和指导。
[引用本文]徐孝东, 梁菊, 刘志红. 溶酶体功能稳态与肾脏疾病[J]. 肾脏病与透析肾移植杂志, 2024, 33(5): 457-462.
XU Xiaodong, LIANG Ju, LIU Zhihong. Lysosomal homeostasis and kidney diseases[J]. Chinese Journal of Nephrology, Dialysis & Transplantation, 2024, 33(5): 457-462.
来源:肾脏病与透析肾移植杂志订阅号
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017