


200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过




摘要:脑膜淋巴管(mLVs)是引流中枢神经系统细胞外液的重要结构,同时也是中枢神经系统的免疫调节器官和保护屏障。近年来,研究者对该结构的解剖、功能、疾病应用等方面的认识逐渐丰富。该文回顾了mLVs的发现历史,阐述了其发生过程和解剖研究的最新进展,重点讨论了mLVs的引流和免疫功能,探讨了脑血管疾病发生发展过程中mLVs的调控机制,以期为脑血管疾病寻找新的治疗策略提供依据。
近年来,大脑缺乏淋巴组织的既往认知逐渐被打破,目前已知的4种淋巴引流途径包括血管周围通路、胶质淋巴、脑膜淋巴管(meningeal lymphatic vessels,mLVs)和嗅-颈引流途径,其中mLVs是将中枢神经系统(central nervous system,CNS)与外周联系起来的重要纽带。
mLVs位于各种脊椎动物的硬膜中,其将CNS的细胞外液(脑脊液、脑间质液)引流到外周的淋巴结。mLVs具有物质引流和平衡脑脊液压力等关键功能,广泛参与蛛网膜下腔出血(SAH)、认知障碍等疾病的病理过程。另外,mLVs参与调节炎性反应和免疫监视,对于神经退行性疾病、癫痫、脑血管疾病、脑肿瘤等具有重要意义。
脑血管病是颅内血液循环障碍所致的神经损伤性疾病,主要分为出血性和缺血性两类。目前,越来越多的研究者关注mLVs与脑血管病之间的关系。本文阐述了mLVs的结构功能,总结了近年来mLVs与脑血管疾病关系的研究进展,并探讨了mLVs调控脑血管疾病的作用机制。
mLVs的发现具有里程碑式的意义。18 世纪,Mascagni和Santi首次发现了硬脑膜中存在“血管淋巴”的组织,但当时并未引起研究者们的重视。Retzius和Key并不认可这一结论,提出了大脑中缺乏淋巴结构的观点。2015年Louveau等和Aspelund等在小鼠硬膜中观察到了广泛分布的淋巴网络,将该结构定义为mLVs,并发现经典的淋巴内皮标志物(包括Prospero同源盒蛋白 1、整合膜蛋白、淋巴管内皮透明质酸受体 1、CC趋化因子配体21和血小板-内皮细胞黏附分子)可以对其进行描绘。
MRI是临床研究观察mLVs引流途径和速度的有效手段。2017年Absinta等利用钆布醇在脑脊液和硬膜之间梯度扩散的原理,使用增强T2液体衰减反转恢复(FLAIR)序列观察了人类和狨猴的mLVs。Kuo等使用无对比剂的冠状时间飞跃序列,显示了人类矢状窦旁mLVs 的流动,Patel等使用7T-MRI对该区域的详细结构进行了研究。Wu等通过三维对比剂增强的T2FLAIR序列观察到百余例可逆性脑血管收缩综合征和丛集性头痛患者的mLVs和胶质淋巴的相互作用。Jacob等通过光片荧光显微镜观察并绘制出了更为详细的人类和小鼠的脑膜淋巴网络。
mLVs是进化保守性器官,研究者们通过观察小鼠和斑马鱼逐渐明晰了mLVs 在个体中的发育过程。小鼠胚胎的mLVs存在于枕骨大孔周围,出生后由两侧翼颚动脉周围开始发生,血管内皮细胞先后表达Prospero同源盒蛋白1和淋巴管内皮透明质酸受体-1,启动淋巴管的分化,在血管内皮生长因子C(vascular endothelial growth factor C,VEGF-C)的作用下mLVs不断发育。mLVs沿着脑膜中动脉向中线延伸,随后延伸到横窦和上矢状窦的两侧,最终到嗅球周围,并在淋巴流动产生的机械力下发育成熟。
虽然斑马鱼与小鼠的大脑结构不同,但mLVs却呈现出类似的生长模式,其mLVs 起源于颅骨形成前两侧的面部淋巴丛,随着颅骨板生长向中线延伸,围绕小脑和视突之间的血管生长。
mLVs根据位置不同可分为背侧颅骨内面和腹侧基底部两部分。淋巴内皮细胞是mLVs的基本结构,其分子特征与外周淋巴管相似,根据有无淋巴瓣膜和平滑肌细胞可以将mLVs分为毛细淋巴管和集合淋巴管。
随着对mLVs的不断研究,其液体引流途径逐步明晰。脑脊液由脑室的脉络丛产生,经过穿动脉的血管周围间隙与大脑间质进行物质交换,最后由蛛网膜颗粒和mLVs排出。蛛网膜下腔的大分子通过mLVs 输送到颈深淋巴结(deep cervical lymph nodes, dCLN)和颈浅淋巴结。虽然血管周围间隙、蛛网膜颗粒和mLVs并无解剖联系,但三者在流体动力学上组成一个整体,相互影响。
近年来,有研究者观察到了mLVs与骨髓之间的解剖联系。Ringstad和Eide通过钆布醇增强MRI观察到脑脊液通过mLVs向骨髓扩散。Mazzitelli等通过向小鼠脑池注射荧光白蛋白也发现了脑脊液沿mLVs向骨髓壁龛内移动的证据。
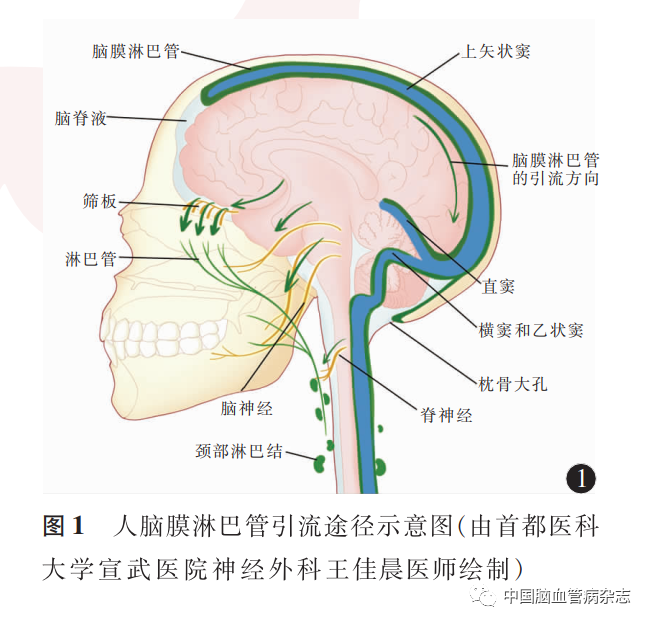
大脑背侧颅内面和腹侧基底的mLVs具有不同的结构和引流功能。背侧颅内面的mLVs主要沿横窦、直窦、乙状窦、后脑静脉、喙鼻静脉以及脑膜中动脉和脑膜前动脉的分支走行,最后沿着枕骨大孔和颈静脉孔流出;腹侧基底部 mLVs沿岩磷窦和乙状窦走行,随后与脑神经伴行,经由神经孔进入外周,继续沿神经外膜的淋巴管进入颈部淋巴结(cervical lymph nodes,CLN)。腹侧基底部 mLVs相比大脑背侧的mLVs更粗大、分支更丰富,有典型的淋巴内皮和瓣膜,毛细淋巴管和集合淋巴管区分明显,因此更适合脑脊液的摄取和引流。腹侧基底部 mLVs在硬膜中的位置靠近蛛网膜下腔,筛板处无蛛网膜分隔,因此能够直接引流脑脊液。mLVs引流途径见图1。
mLVs作为CNS与外周免疫的桥梁,可以转运硬膜中的免疫细胞,调节神经炎性反应。T淋巴细胞获取CNS 抗原是神经炎性反应的关键过程。在实验性自身免疫性脑脊髓炎小鼠模型中,mLVs内皮细胞上调与白细胞黏附趋化和活化调节相关基因的表达,增加与细胞黏附分子11c结合的能力,并以干扰素γ依赖的形式,上调Ⅱ类主要组织相容性复合体和程序性死亡受体配体1的表达,增加了与分化簇4阳性T淋巴细胞和树突状细胞的相互作用,从而促进抗原呈递,实现适应性免疫调节。mLVs周围的树突状细胞可以限制滤泡辅助T淋巴细胞并调节T淋巴细胞,从而抑制CNS免疫。mLVs周围的B淋巴细胞活化与脊髓病变的程度相关。颅内感染、肝性脑病等疾病通过VEGF-C诱导mLVs生成,新生的mLVs 增加了对炎性免疫细胞的输入和输出。
当mLVs消融后,日本脑炎病毒感染的小鼠大脑中的中性粒细胞、巨噬细胞、单核细胞、分化簇8阳性T淋巴细胞、分化簇4阳性T淋巴细胞、NK细胞、小胶质细胞和炎性细胞因子(白细胞介素6、肿瘤坏死因子α、白细胞介素1b)显著增加,对神经元造成损伤。消融mLVs的阿尔兹海默病小鼠模型表现出明显的淀粉样β蛋白沉积,小胶质细胞增生和神经血管障碍,突触可塑性和神经传递的转录改变,表现为学习和记忆功能缺陷。
mLVs是CNS免疫屏障的一部分,其通过不断感知血液和脑脊液中成分的变化,发挥免疫监视作用。Hu等通过胶质瘤和黑色素瘤脑转移小鼠模型发现,VEGF-C通过C-C 趋化因子受体7及其配体21途径显著增强程序性死亡受体1抗体的抗肿瘤疗效,并证明其机制为VEGF-C通过促进mLVs增生,增加了肿瘤中细胞毒性T淋巴细胞和树突状细胞的浸润,从而强化了细胞免疫监视脑组织的功能。mLVs还能通过清除衰老的细胞来发挥免疫监视作用。衰老的星形胶质细胞高表达VEGF-C,通过C-C趋化因子受体7途径经由mLVs被清除。因此或可通过增加VEGF-C的表达水平以强化mLVs的免疫监视功能,使大脑在衰老过程中维持稳态。
脑血管疾病主要包括出血性和缺血性两类,二者具有类似的脑脊液循环和神经炎性反应机制。研究mLVs在脑血管病中的作用有助于深入了解疾病的发生机制并提供潜在的治疗策略。
5.1.1 SAH:Pu等向小鼠脑脊液中注射红细胞以模拟SAH后观察到mLVs和dCLN中汇集了大量红细胞,表明mLVs是积存红细胞的途径之一。Chen等使用光敏感剂维替泊芬激光消融SAH模型小鼠的mLVs或使用化学抑制剂MAZ51竞争性抑制VEGF受体3活性后,dCLN的红细胞数量显著减少,小胶质细胞比例显著增加,神经炎性反应加重,导致神经损伤、行为缺陷和记忆障碍加重。SAH发生后血凝块可能阻塞mLVs,影响引流功能,使无法排出的脑脊液进入血管周围间隙,损伤水通道蛋白4,导致毒性和炎性代谢物过分堆积,继而可能诱发长期的脑损伤。而mLVs能高效清除出血,在红细胞裂解为血红蛋白前将其排出蛛网膜下腔,从而有效避免血红蛋白暴露引发的炎性反应。
Liao等发现,酮洛芬、9-顺式维甲酸和VEGF-C可以维持mLVs的完整性并改善引流功能,促进出血后的脑水肿吸收。光生物调节是一种增强mLVs引流的方法,Salehpour等通过光生物调节显著增强了mLVs对红细胞的清除能力,该方法通过线粒体细胞色素C氧化酶影响淋巴内皮细胞的氧化还原电位,进而调节mLVs的通透性和延展性。上述结论均来自于啮齿动物实验,人类SAH发生时mLVs的功能状态仍待更多探索。
5.1.2硬膜下血肿(subdural hematoma,SDH):SDH是蛛网膜和硬脑膜之间的血肿,该血肿一定程度上能够自发吸收,但引流途径尚不明确。Liu等证明了SDH的mLVs引流机制,他们向小鼠硬膜下间隙注入血液、纤维蛋白原和荧光标记葡萄糖,示踪剂沿着dCLN中的淋巴内皮标志物(即淋巴管内皮透明质酸受体1)阳性的淋巴窦流动,6h后红细胞聚集于dCLN中。SDH下调淋巴管内皮透明质酸受体1和VEGF-C的表达水平,导致mLVs引流能力下降。颅颈人工淋巴引流作为一种理疗方法,能够提高药物治疗慢性SDH 的疗效,促进血肿的清除,该方法耐受性好,易于操作,在其他脑血管病中的效果有待进一步验证。
5.1.3脑实质出血(intracerebral hemorrhage,ICH):mLVs能够有效清除脑实质内的血肿占位。Semyachkina-Glushkovskaya等观察到ICH小鼠早期mLVs引流增强,dCLNs中含铁血黄素迅速聚集,ICH发生1h后 mLVs直径扩大了3~4倍。Tsai等[38]发现,ICH小鼠晚期mLVs增生,ICH 发生后第14天mLVs的新生分支和表面积增大,对示踪剂的清除能力增强,大脑背侧mLVs更为显著,可以持续至少60d。该研究发现,磷酸二酯酶抑制剂西洛他唑可以改善mLVs功能,增加对红细胞的摄取和清除,减轻铁沉积、神经元变性凋亡和胶质细胞增多所带来的不良后果,有效改善预后,但具体的作用机制尚不明确。
CIS是致人死亡和残疾的严重疾病,CIS所致的急性脑水肿是预后不良的重要因素。Yanev等将小鼠的脑膜小血管栓塞2周后发现,脑膜缺血刺激外侧矢状窦附近的mLVs增生,而栓塞大脑中动脉不能诱导mLVs增生,这表明与mLVs解剖关系接近的缺血更容易刺激mLVs的新生,同时通过三维序列双光子断层扫描成像技术对脑梗死面积和脑室体积进行观察发现,缺血部位周围的脑组织水肿也得到减轻。
mLVs不仅能减轻CIS所致的脑水肿,还能诱导血管再生。Chen等通过斑马鱼CIS模型发现,mLVs向受损的脑实质生长并形成腔体,诱导血管生成,新生血管成熟后,淋巴腔体凋亡消失,恢复大脑实质的无淋巴状态。两年后该研究团队更新了研究结论,认为CIS后新生淋巴管可以直接转分化为血管,其机制与促红细胞生成素生成肝癌相互作用体B2a及Notch信号通路调节有关。新生淋巴管的生长方向依赖于残余血管释放的基质衍生因子-1b的引导。目前相关研究的对象仅为斑马鱼,尚待在其他实验动物中验证。促进血管再生是未来治疗缺血性卒中的方向,继续研究mLVs 诱导血管再生的作用具有重要意义。
脑血管栓塞后神经元损伤可引起广泛的炎性反应。Esposito等发现,小鼠大脑中动脉栓塞72h内,CLN中巨噬细胞被迅速激活,手术切除CLN或化学阻断VEGF受体3可减轻炎性反应,缩小梗死区域体积。该研究结果表明,CLN参与卒中后的炎性反应调节,但仍有待进一步证明其与mLVs调节之间的联系。Bai等应用颅骨搬移手术改善了大脑中动脉闭塞小鼠mLVs的引流功能,手术后mLVs和dCLNs摄取的荧光标记白蛋白明显增加,缺血区域T淋巴细胞浸润明显减少。探索增强mLVs引流功能的外科手段具有积极意义,可作为靶向治疗缺血性卒中的潜在方向。
mLVs是位于硬脑膜中的解剖结构,在脑血管疾病中体现出一定的适应性和代偿能力,能够及时将出血或积液引流至CLN,调节炎性反应,保护神经系统。目前针对mLVs的研究方法主要集中于动物模型的免疫化学染色和显微镜观察,临床数据尚待完善。MRI可实现mLVs的临床观察,目前已应用于多发性硬化、帕金森综合征、阿尔兹海默病和脑肿瘤等疾病的mLVs评估,但尚未在脑血管病患者中开展。增强mLVs引流和免疫调节功能可以改善多种脑血管病动物模型的预后,其方法包括化学或理学方法,例如病毒载体导入VEGF-C、颅颈淋巴引流和光生物调节,也可以通过外科手术进行干预。
未来研究的方向或将聚焦于以下几点:(1)明确mLVs对出血和缺血性脑血管疾病引发的脑水肿的引流调节机制,以便有效清除占位和控制炎性反应。(2)增强外周淋巴功能的药物对mLVs的有效性和可靠性有待进一步验证,探索物理学和外科手段改善淋巴功能具有临床研究价值。(3)目前的研究高度依赖动物模型,因此收集更多临床标本和数据有助于深入认识人类mLVs的作用机制。
本文转自:中国脑血管病杂志
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017